"Sleep is regulated by two distinct biological drives: Process S (the homeostatic sleep pressure generated by adenosine accumulation) and Process C (the circadian rhythm driven by the suprachiasmatic nucleus). Mastering your sleep requires aligning these two forces to achieve a perfect intersection at bedtime."
Key Takeaways: Sleep Pressure vs. Circadian Rhythm
-
1.
Process S (Sleep Pressure): An accumulation of the inhibitory neurotransmitter adenosine in the basal forebrain, which builds continuously while awake and clears during deep sleep.
-
2.
Process C (Circadian Rhythm): An internal 24-hour clock driven by the Suprachiasmatic Nucleus (SCN) that controls melatonin, cortisol, and core body temperature cycles independently of sleep duration.
-
3.
The Dual-Process Alignment: Optimal sleep occurs when sleep pressure is at its peak (high adenosine) and the circadian arousal signal declines (low cortisol, rising melatonin).
Introduction: The Two-Process Model of Sleep Regulation
Why do we sleep when we do? And why is it that sometimes, even when we are completely exhausted after a long day of travel, we still struggle to fall asleep or wake up after just a few hours? The answers lie in the two-process model of sleep regulation, first proposed in 1982 by Swiss sleep scientist Dr. Alexander Borbély. This landmark model describes sleep as the result of a dynamic interaction between two distinct physiological processes: Process S, the homeostatic sleep drive, and Process C, the circadian rhythm.
Many biohackers focus heavily on optimizing their circadian rhythm through morning sunlight and blue-blocking glasses. While essential, this only addresses one half of the sleep equation. If you have not built up sufficient sleep pressure throughout the day, or if you have inadvertently cleared your sleep debt prematurely, circadian alignment alone will not rescue your sleep latency or depth. Conversely, high sleep pressure in a misaligned circadian state leads to fragmented, low-quality rest. For extreme longevity and cognitive preservation, you must learn to measure, balance, and align both forces.
Process S: The Biochemical Basis of Sleep Pressure
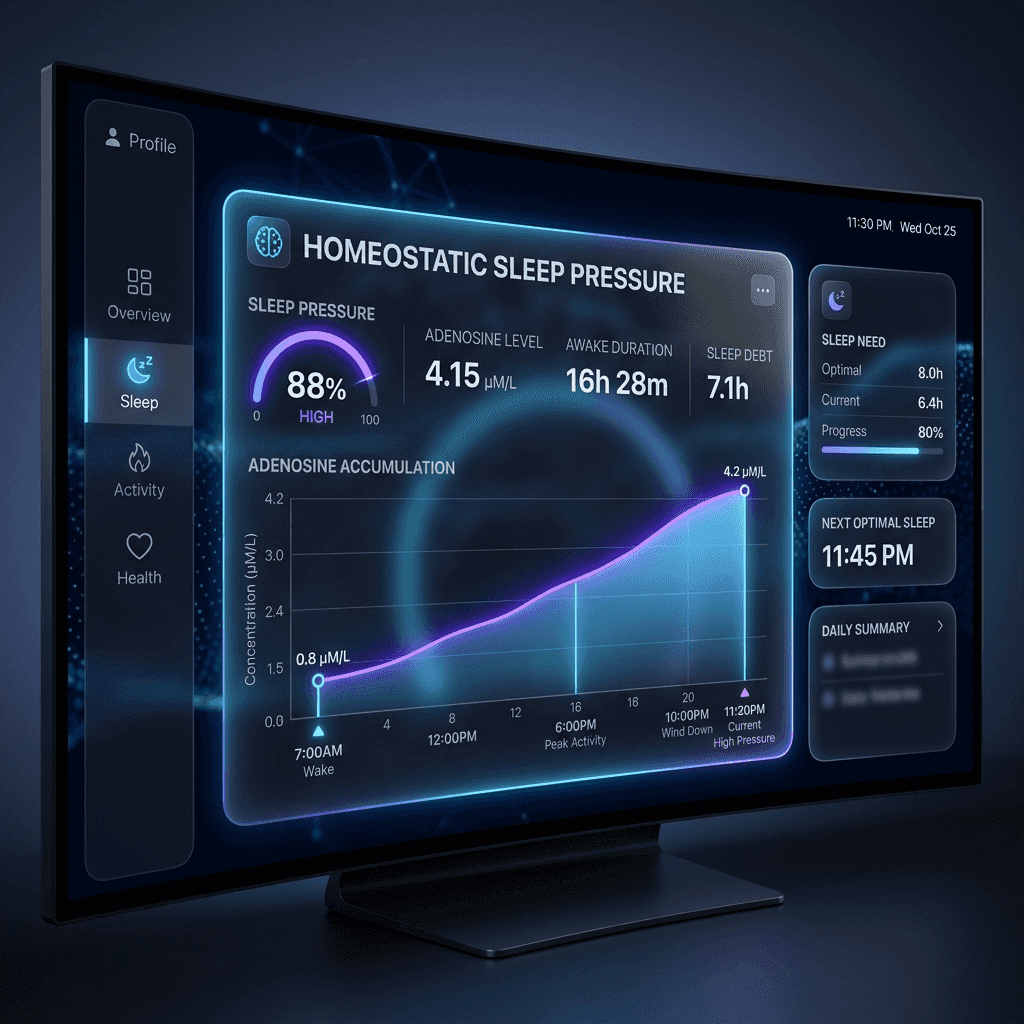
Process S is a neurochemical sleep debt. Every minute you are awake, your brain cells consume energy in the form of Adenosine Triphosphate (ATP). As ATP is metabolized to support cellular function, it breaks down into adenosine. This adenosine slowly accumulates in the extracellular spaces of the basal forebrain. As adenosine levels rise, it binds to specific adenosine A1 and A2A receptors on sleep-promoting and wake-promoting neurons.
The binding of adenosine to A1 receptors suppresses the activity of cholinergic, adrenergic, and histaminergic arousal centers in the brainstem, while binding to A2A receptors actively excites sleep-promoting neurons in the ventrolateral preoptic nucleus (VLPO). The result is a progressive sensation of sleepiness—what we refer to as sleep pressure or sleep drive. The only way to clear adenosine from these receptors is through slow-wave sleep. During deep sleep, the brain metabolizes and flushes away accumulated adenosine, resetting your sleep debt to zero for the next waking cycle.
Biohacker Pro-Tip: The 90-Minute Caffeine Delay
Caffeine is an adenosine receptor antagonist; it blocks adenosine from binding to A1 and A2A receptors without activating them, temporarily masking sleep pressure. If you consume caffeine immediately upon waking, you block the remaining residual adenosine before it has cleared. Once the caffeine metabolizes, the sudden rush of accumulated adenosine triggers a severe afternoon crash. Delay your first cup of coffee by 90 to 120 minutes after waking. This allows your body to naturally clear the last of your overnight adenosine, stabilizing your energy levels and maintaining high evening sleep pressure.
The Biophysics of Adenosine Receptors & Caffeine Interference
To master Process S, we must look at the molecular level. Adenosine is a purine nucleoside. The adenosine A1 receptor is coupled to Gi proteins, meaning that when adenosine binds, it inhibits adenylyl cyclase, reducing intracellular cyclic AMP (cAMP) and closing calcium channels. This suppresses the release of excitatory neurotransmitters like glutamate. In contrast, the A2A receptor is coupled to Gs proteins, which stimulates adenylyl cyclase, raising cAMP and exciting GABAergic sleep-promoting neurons within the VLPO.
Caffeine has an almost identical molecular structure to adenosine. This structural similarity allows caffeine to fit perfectly into the binding pockets of both A1 and A2A receptors. By blocking these sites, caffeine acts as a competitive antagonist, preventing endogenous adenosine from signaling sleepiness. However, while the perception of sleep pressure is masked, the production of adenosine does not stop. It continues to pile up in the synapse. When the liver's cytochrome P450 enzymes finally break down the caffeine, the accumulated adenosine floods the receptors all at once, triggering a severe energy crash. This biochemical reality is why caffeine timing is one of the most critical levers for stabilizing sleep architecture.
Process C: The Circadian Pacemaker
Process C is a 24-hour cycle that operates independently of how long you have been awake. Even if you stay awake for 48 hours, Process C will continue to signal alertness during the day and sleepiness at night. This rhythm is generated by the master circadian clock: the Suprachiasmatic Nucleus (SCN), a tiny cluster of about 20,000 neurons located in the hypothalamus, directly above the optic chiasm.
The SCN receives direct light information from the eyes via the retinohypothalamic tract. Sensory-evoked inputs from intrinsically photosensitive retinal ganglion cells (ipRGCs) detect the presence of short-wavelength blue light (approx. 480 nm), signaling the start of the day. In response, the SCN stimulates the adrenal glands to release a cortisol pulse, raising heart rate, blood pressure, and core body temperature. As daylight fades, the lack of blue light tells the SCN to signal the pineal gland to synthesize melatonin, which lowers core body temperature and prepares the body for sleep.
The SCN Molecular Loop (CLOCK/BMAL1/PER/CRY)
At the core of SCN cells is a genetic clock mechanism: the Transcription-Translation Feedback Loop (TTFL). This loop operates on a 24.2-hour cycle and is governed by four primary clock genes: CLOCK, BMAL1, Period (PER), and Cryptochrome (CRY). During the morning, the CLOCK and BMAL1 proteins bind together in the cell nucleus, acting as transcription factors that activate the transcription of the PER and CRY genes.
As PER and CRY proteins accumulate in the cytoplasm, they form complexes and migrate back into the nucleus. Once inside the nucleus, they bind to the CLOCK-BMAL1 complexes, suppressing their own transcription. Over the course of the night, the PER and CRY proteins are slowly degraded. By morning, the suppression is lifted, allowing the cycle to start again. This intracellular genetic loop acts as the primary clock driver for every cell in the human body, coordinating tissue-specific processes like liver metabolism, muscle repair, and immune activation.
Process S vs. Process C Comparison
| Characteristic | Process S (Sleep Pressure) | Process C (Circadian Rhythm) |
|---|---|---|
| Biological Nature | Homeostatic & Accumulative | Oscillatory & Rhythmic |
| Primary Biomarker | Extracellular Adenosine in basal forebrain | Melatonin, Cortisol, Core body temperature |
| Control Center | Basal Forebrain & VLPO Nucleus | Suprachiasmatic Nucleus (SCN) in Hypothalamus |
| Influenced By | Waking hours, physical activity, caffeine intake | Light exposure (blue spectrum), feeding cycles, social cues |
| Resolution Mode | Slow-wave deep sleep (N3) duration | 24-hour rotation of light and dark states |
Peripheral Cellular Clocks & Nutrient-Sensing Pacemakers
While the SCN is the master pacemaker, it does not act alone. Every peripheral organ (liver, gut, pancreas, skeletal muscle) contains its own cell-autonomous clock governed by the same TTFL loops. These peripheral clocks are synchronized by the SCN but are also highly sensitive to non-photic cues, particularly nutrient intake. When we eat, the pancreas releases insulin and the liver processes nutrients, signaling wakefulness and digestion to these peripheral clocks.
If you eat a large meal in the middle of the night, your master SCN clock says "it is dark, sleep," but your liver and gut clocks say "it is light, digest." This state of internal desynchronization disrupts metabolic pathways, impairs insulin sensitivity, and causes systemic inflammation. To maintain cellular alignment, you must use both light (for the SCN) and food intake (for peripheral clocks) as coordinated anchors.
Three Protocols to Align Process S and Process C
Morning Bright Light Anchoring (Process C)
To align your SCN, expose your eyes to bright light within 30 minutes of waking. Natural sunlight is the single most powerful circadian cue (zeitgeber). On a clear day, outdoor light provides 10,000 to 100,000 lux, whereas typical indoor lights only provide 100 to 500 lux. In winter or dark climates, use a specialized 10,000-lux light box or light therapy glasses (like Luminette).
This morning light anchor shuts down melatonin production instantly and programs your pineal gland to start melatonin synthesis exactly 14 to 16 hours later, aligning Process C to a predictable evening schedule.
Physical Exercise & Adenosine Maximization (Process S)
Because sleep pressure is generated by cellular energy expenditure, physical activity is the most effective way to accelerate adenosine accumulation. Engaging in high-intensity cardiovascular training or heavy resistance training significantly speeds up the breakdown of ATP, building a steep sleep pressure gradient.
To protect sleep architecture, perform intense exercise in the morning or early afternoon. Exercising within 4 hours of bedtime raises your core body temperature and cortisol levels, which conflicts with Process C's cooling requirements and delays melatonin release.
The 10-Hour Caffeine & 3-Hour Feeding Curfews
To prevent interference with your homeostatic and circadian drives, implement strict temporal curfews. Caffeine has a half-life of 5 to 7 hours and a quarter-life of up to 12 hours. Consuming caffeine past 12:00 PM means a significant amount remains in your brain at 10:00 PM, blocking adenosine receptors and preventing deep slow-wave sleep.
Similarly, stop eating all food at least 3 hours before sleep. Digestion raises core body temperature and triggers insulin release, which signals wakefulness to peripheral cellular clocks in your organs, disrupting SCN alignment and fragmented sleep quality.
The Role of Peripheral Food & Temperature Clocks
Although the suprachiasmatic nucleus (SCN) acts as the master conductor, every tissue and organ in the body contains its own peripheral clock. These peripheral clocks (in the liver, pancreas, gut, and adipose tissue) regulate local functions such as nutrient absorption, glucose homeostasis, lipid metabolism, and bile acid synthesis. While the SCN is primarily entrained by light-dark cycles, peripheral clocks are highly responsive to nutrient availability and metabolic signals. Eating acts as a powerful non-photic synchronizer (zeitgeber) for these metabolic tissues.
When you consume food, digestion triggers the release of insulin, which directly upregulates the transcription of core clock genes in the liver. If you eat during your biological night (when the SCN is signaling rest), your liver clock shifts out-of-phase with the SCN. This desynchronization leads to a state of metabolic chaos: liver glycogen synthesis is disrupted, insulin sensitivity drops, and systemic lipid metabolism is impaired, driving abdominal fat accumulation and metabolic syndrome. To maintain systemic alignment, you must restrict your eating window to daylight hours, ensuring that food signals match the SCN's light-induced alertness phase.
Non-Photic Zeitgebers: Timing Exercise & Social Cycles
Beyond light and food, physical activity and social interactions act as secondary non-photic zeitgebers. Skeletal muscle cells contain autonomous clocks that regulate mitochondrial respiration, glucose uptake, and protein synthesis. Engaging in intense physical exercise triggers a cascade of intracellular signals, including AMPK activation and calcium-calmodulin-dependent kinase signaling, which reset the muscular clock and influence the central SCN pacemaker via systemic temperature shifts and hormonal pathways.
The timing of exercise is therefore critical for circadian phase adjustment. Morning exercise (between 7:00 AM and 10:00 AM) promotes a phase advance, making it easier to fall asleep earlier in the evening. Late-evening exercise (after 8:00 PM), however, causes a phase delay. The sudden spike in core body temperature and cortisol levels conflicts with the SCN's natural evening cooling cycle, delaying melatonin synthesis and disrupting sleep architecture. To optimize both sleep pressure and circadian rhythm, focus your high-intensity workouts during the morning or early afternoon, leaving your evenings for low-intensity somatic stretching and relaxation.
Age-Related Desynchronization of the Dual Drives
As we navigate the biological trajectory of aging, the coordination between Process S and Process C begins to deteriorate. On one side of the equation, the decline in slow-wave sleep amplitude (a symptom of aging prefrontal cortex tissues) prevents the complete overnight clearance of adenosine. As a result, older individuals often wake up with high levels of residual adenosine, meaning their starting sleep pressure debt is never fully reset to zero. This leads to persistent daytime drowsiness and a blunted sleep pressure gradient throughout the day, which makes it harder to trigger deep slow oscillations on the subsequent night.
On the other side of the equation, the suprachiasmatic nucleus (SCN) undergoes structural aging. The master clock suffers from a loss of synaptic plasticity, and the pineal gland often calcifies, significantly blunting the evening melatonin release. This circadian flattening reduces the strength of the core body temperature decline, resulting in fragmented sleep and frequent nighttime awakenings. To counteract this age-related desynchronization, older adults must pay close attention to light hygiene and non-photic zeitgeber anchors. Incorporating closed-loop acoustic stimulation (to amplify delta power) and utilizing micro-dose exogenous melatonin therapy (0.3 mg taken 2 hours before bed) can help restore the alignment of these dual drives, preserving deep recovery phases as we age.
Furthermore, researchers have mapped the genetic variations that influence SCN resilience during aging. Polymorphisms in clock genes like *PER3* can dictate whether an individual is a "morning larks" or a "night owls" and how severely their circadian pacemaker degrades over time. By utilizing continuous temperature tracking (via smart rings or skin sensors) to map your core body temperature minimum (which typically occurs 2 hours before waking), you can identify your exact circadian phase position and adjust your light exposure and eating windows to prevent age-related drift, keeping your internal clock highly synchronized. Specifically, mutations in the *CRY1* gene have been linked to Delayed Sleep Phase Disorder (DSPD), a common condition where the internal circadian cycle is stretched, causing individuals to sleep and wake significantly later. By understanding these genetic baseline chronotypes, biohackers can tailor their morning light therapy lux levels and evening melatonin micro-doses with surgical precision, forcing a synchronization of their peripheral and central clocks despite genetic predispositions.
The Clinical Pathology of Desynchronization
What are the physiological consequences of chronic desynchronization? When Process S and Process C are chronically misaligned—such as in shift workers or individuals with severe social jet lag—the body remains in a state of persistent physiological stress. The constant mismatch between central and peripheral cellular clocks prevents the normal overnight downregulation of inflammatory pathways, driving chronic low-grade inflammation (inflammaging).
This desynchronization also disrupts metabolic pathways, leading to impaired glucose tolerance, increased visceral fat storage, and elevated cardiovascular risks. By prioritizing the structural alignment of light exposure, physical activity, and nutrient timing, you protect your body from the systemic wear and tear of circadian disruption, extending biological healthspan.
Peer-Reviewed Clinical Validations & Extended Deeper Reading:
- The Two-Process Model Proposal: Borbély (1982). "A two-process model of sleep regulation". Human Neurobiology. The original mathematical and conceptual framework detailing the interaction between Process S and Process C. Read Two-Process Study
- Adenosine & Basal Forebrain: Porkka-Heiskanen et al. (1997). "Adenosine: A mediator of the sleep-inducing effects of prolonged wakefulness". Science. Clinical validation that sleep deprivation increases extracellular adenosine concentrations in sleep-regulating brain structures. Read Adenosine Study
- SCN Master Clock & ipRGCs: Berson et al. (2002). "Phototransduction by retinal ganglion cells that project to the suprachiasmatic nucleus". Science. Describes the discovery of the specialized blue-light-sensitive cells in the retina that synchronize the circadian master pacemaker. Read Retinal Ganglion Study